3.1. Alpha Helix adalah sebuah struktur koil
tersetabilkan oleh ikatan hidrogen antar rantai.
Di dalam mengevaluasi struktur potensial,
Pauling dan Corey mempertimbangkan penyesuaian sterik peptida yang memungkinkan
dan yang paling secara penuh memanfaatkan kapasitas ikatan hidrogen rangka
gugus NH dan CO. Struktur pertama yang diusulkan mereka, adalah suatu heliks,
yang mana bentuk struktur seperti tangkai (Gambar. 29). Suatu rangka bergulung
membentuk bagian dalam tangkai dan rantai sisi meluas keluar seperti suatu
pilinan. Suatu heliks distabilkan oleh ikatan hidrogen antara gugus NH dan CO
rantai utama. gugus CO masing-masing bentuk asam amino adalah suatu ikatan
hidrogen dengan gugus NH asam amino yang diposisikan pada empat residu (Gambar.
30). Kecuali asam amino yang bagian ujung dari suatu suatu heliks, gugus CO dan
NH rantai utama semua memungkinkan berikatan hidrogen. Masing-Masing residu
dihubungkan dengan satu yang berikutnya oleh suatu kenaikan sepanjang poros
garis heliks 1.5Å dan suatu putaran 100 derajat, yang memberi 3.6 amino residu
asam tiap putaran heliks.
Seperti itu, amino yang asam spasi tiga
dan empat terpisah di dalam urutan dengan leluasa dan dekat satu sama lain di
dalam suatu suatu garis heliks. Di dalam kontras dua asam, amino yang terpisah
dalam urutan diposisikan pada sisi berlawanan heliks dan sehingga tidak terjadi
kontak. Lilitan suatu heliks adalah sepadan dengan produk terjemahan (1.5Å) dan
banyaknya residu tiap putaran (3.6), 5.4Å. Pependekatanan/Pengertian Sekrup
suatu heliks dapat menarah kanan (searah jarum jam) atau mengarah kiri (berlawanan
arah jarum jam). Diagram Ramachandran mengungkapkan kedua-duanya (kanan dan
kiri) yang memungkankan terjadi (Gambar. 31). Bagaimanapun, heliks kanan dengan
lebih baik sebab lebih sedikit halangan sterik antara rantai sisi dan rantai
induk. Yang sangat utama semua α-heliks yang ditemukan di dalam) protein adalah
putaran kiri.
Di dalam diagram protein menurut bagan, α-
helices dilukiskan seperti terbelit pita atau tangkai (Gambar. 32).

Gambar. 29. Struktur suatuheliks. (A)
Suatu lukisan pita dengan atom α-karbon dan rantai sisi (hijau). (B) Tampak
samping suatu versi bola dan tangkai melukiskan ikatan hidrogen (garis
terputus) antara gugus NH dan CO. (C) Suatu tampilan akhir menunjukkan rangka yang
bergulung ketika bagian dalam heliks dan rantai sisi (hijau) diproyeksian
keluar. (D) Suatu tampolan space-filling bagian C menunjukkan inti bagian dalam
yang dengan ketat memenuhi heliks.

Gambar.30. Rancangan ikatan jidrogen
Karena suatu heliks. Di dalam suatu heliks, gugus CO residu n membentuk suatu
ikatan hidrogen dengan gugus NH residu n +4.

Pendekatan Skrup
Menjelaskan arah di mana struktur heliks berputar terhadap
porosnya. Jika, dilihat bawah sumbu heliks, rantai berubah searah jarum jam, ia
memiliki pendekatan sekrup kanan. Jika memutar berlawanan adalah, pengertian
sekrup kidal. Pauling dan Corey meramalkan struktur heliks 6 tahun sebelum
benar-benar melihat dalam rekonstruksi x-ray dari struktur mioglobin.
Penjelasan dari struktur heliks adalah tengarai/diperkirakan dalam biokimia
karena menunjukkan bahwa konformasi dari rantai polipeptida dapat diprediksi
jika sifat komponennya secara ketat dan tepat telah dikenali.
kisaran prosentase isi α-heliks protein sangat luas, dari
hampir tidak ada sampai hampir 100%. Sebagai contoh, sekitar 75% dari residu
dalam feritin, yakni protein yang membantu menyimpan zat besi, berada dalam
heliks (Gambar. 33). Heliks tunggal biasanya kurang dari 45Ǻ panjang. Namun,
dua atau lebih merupakan heliks yang dapat terbelit untuk membentuk struktur
yang sangat stabil, dengan panjang dapat mencapai 1000 Å (100 nm, atau 0,1 mm)
atau lebih (Gambar. 34). Seperti koil-heliks melingkar ditemukan di myosin dan
tropomyosin dalam otot, di fibrin dalam pembekuan darah, dan di keratin dalam rambut.
Untaian heliks dalam protein melayani peran mekanik dalam membentuk bundel yang
kaku dari serat, seperti pada duri landak. Sitoskeleton (perancah internal) sel
banyak mengandung sesuatu yang disebut filamen menengah, yang juga dua koil-α-heliks
beruntai yang tergulung. Banyak protein yang membran span biologis juga
mengandung heliks.


Figure. 32. Skema α-Helices. (A) Sebuah model tangkai dan bola. (B)gambaran
piata. (C) gambaran tabung.

Gambar. 33. α-heliks protein besar. Feritin,
protein penyimpan-besi, dibangun
dari bundel a-heliks.

Gambar. 34. Sebuah
Coil a-spiral melingkar. Kedua heliks
memutar di sekitar satu
sama lain untuk membentuk suatu
superhelix. Struktur seperti ini ditemukan dalam banyak protein termasuk keratin pada rambut, cakar, dan tanduk.
3.2.
Beta Sheets distabilkan oleh Ikatan Hidrogen Antara Strands Polipeptida
Pauling dan Corey menemukan motif lain struktur periodik, yang mereka namakan β-pleated sheets (β karena itu struktur yang kedua dijelaskan, setelah helix yang telah pertama). β-pleated sheets (atau, lebih sederhana, β-sheets) berbeda nyata dari suatu helix rodlike. Sebuah rantai polipeptida, yang disebut untai ab, dalam sheets ab hampir sepenuhnya diperpanjang bukannya erat melingkar seperti dalam heliks. Berbagai struktur diperpanjang sterik diperbolehkan (Gambar. 35).
Pauling dan Corey menemukan motif lain struktur periodik, yang mereka namakan β-pleated sheets (β karena itu struktur yang kedua dijelaskan, setelah helix yang telah pertama). β-pleated sheets (atau, lebih sederhana, β-sheets) berbeda nyata dari suatu helix rodlike. Sebuah rantai polipeptida, yang disebut untai ab, dalam sheets ab hampir sepenuhnya diperpanjang bukannya erat melingkar seperti dalam heliks. Berbagai struktur diperpanjang sterik diperbolehkan (Gambar. 35).

Gambar . 35. Ramachandran
Diagram Untuk b Strands.
Area merah menunjukkan konformasi sterik yang diperbolehkan diperpanjang, b-
Strands -seperti struktur.
Jarak antara asam amino yang berdekatan di sepanjang untai
ab adalah sekitar 3,5 Å, berbeda dengan jarak 1,5 Å sepanjang heliks. Rantai
samping asam titik yang berdekatan amino dalam arah yang berlawanan (Gambar. 36).
Sebuah β sheets dibentuk dengan menghubungkan dua atau lebih helai β oleh
ikatan hidrogen. Rantai yang berdekatan di αβ-sheets dapat berjalan di arah
yang berlawanan (antiparalel α-sheets) atau di arah yang sama (paralel β-sheets).
Dalam pengaturan antiparalel, gugus NH dan CO gugus masing-masing asam amino
yang masing-masing hidrogen terikat pada gugus CO dan gugus NH dari mitra pada rantai
yang berdekatan (Gambar. 37). Dalam susunan paralel, skema ikatan hidrogen
sedikit lebih rumit. Untuk setiap asam amino, gugus NH adalah hidrogen terikat
pada gugus CO dari satu asam amino pada untai yang berdekatan, sedangkan gugus
CO adalah hidrogen terikat pada gugus NH pada asam amino dua residu jauh di
sepanjang rantai (Gambar. 38). Banyak helai, biasanya 4 atau 5, tetapi isa juga
bsebanyak 10 atau lebih, dapat datang bersama-sama dalam β-sheets. β-sheets
tersebut dapat murni antiparalel, murni paralel, atau campuran (Gambar. 39).
Dalam diagram skematis, b untaian biasanya digambarkan oleh anak panah yang
luas menunjuk ke arah ujung karboksil-terminal untuk menunjukkan jenis β-sheets
paralel atau antiparalel yang terbentuk. Lebih struktural beragam dari heliks, β-sheets
dapat relatif datar tetapi kebanyakan mengadopsi bentuk agak bengkok (Gambar, 40).
β-sheets adalah elemen struktural penting dalam banyak protein. Sebagai contoh,
asam lemak-mengikat protein, penting untuk metabolisme lemak, yang dibangun
hampir seluruhnya dari β-sheets (Gambar. 41).

Gambar .36. Struktur dari b-Strand . Rantai samping (hijau) secara bergantian di atas dan di
bawah bidang strand tersebut.

Gambar. 37. Sebuah antiparalel b-Sheet. Berdekatan b-strands helai berjalan di arah yang
berlawanan. Ikatan Hidrogen antara gugus CO dan NH
menghubungkan masing-masing asam amino ke asam amino
tunggal pada strands yang berdekatan, serta menstabilkan struktur.

Gambar. 38. Sebuah Paralel bsheet. Berdekatan b sheet berjalan
dalam arah yang sama. Ikatan Hidrogen menghubungkan masing-masing asam amino pada satu strand dengan dua asam amino yang
berbeda pada strand berdekatan.

Gambar. 39. Struktur Campuran b-sheet .
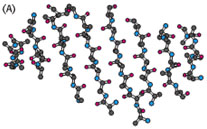

Gambar. 40. Sebuah twisted b sheet. (A) Sebuah model bola-dan-tongkat. (B) Sebuah model
skema. (C) Pandangan skematik diputar 90 derajat untuk menggambarkan lebih
jelas bentuk
memelintirnya.

Gambar. 41. Sebuah Protein yang
banyak b sheet. Struktur asam
lemak mengikat protein.
3.3.
Rantai polipeptida Dapat Mengubah Arah oleh Pembentukan Reverse Turns dan Loops
Protein memiliki kekompakan, bentuk bulat, yang memerlukan
pembalikan ke arah rantai polipeptidanya. Banyak dari pembalikan yang dilakukan
oleh elemen struktural umum disebut reverse
turn (juga
dikenal sebagai β turn atau hairpin
βend), diilustrasikan pada Gambar. 42.
Dalam banyak hal berubah sebaliknya, gugus CO dari residu 1 dari polipeptida
adalah hidrogen terikat pada gugus NH residu 1 + 3. Interaksi ini menstabilkan
perubahan mendadak dalam arah rantai polipeptida.

Gambar. 42. Struktur Turn
Lookup. Kelompok CO dari residu i dari rantai polipeptida berikatan hidrogen pada gugus NH
residu i + 3 untuk menstabilkan turn/belokan.
Dalam kasus lain, struktur yang lebih rumit yang bertanggung
jawab untuk pembalikan rantai. Struktur ini disebut loop atau kadang-kadang ω-loop
(loop omega) untuk menunjukkan bentuk keseluruhan mereka. Tidak seperti heliks
dan untai-β, loop tidak memiliki keteraturan, struktur periodik. Meskipun
demikian, struktur loop sering kaku dan dpat didefinisikan dengan baik (Gambar.
43). Ternyata dan loop selalu berbaring pada permukaan protein dan dengan
demikian sering berpartisipasi dalam interaksi antara protein dan molekul
lainnya. Distribusi dari heliks, β-untai, dan ternyata sepanjang rantai protein
sering disebut sebagai struktur sekunder.

Gambar. 43. Loop pada Permukaan
Protein. Sebuah bagian dari sebuah antibodi molekul memiliki permukaan loop (ditampilkan
dalam warna merah) yang memediasi interaksi dengan molekul lain.
3.4.
Struktur tersier: Gelombang Lipatan Protein Ke Struktur kompak dengan Cores nonpolar
Mioglobin, pembawa oksigen dalam otot, merupakan rantai
polipeptida tunggal 153 asam amino. Kapasitas mioglobin untuk mengikat oksigen
tergantung pada adanya heme, suatu gugus prostetik nonpolypeptide (pembantu)
yang terdiri dari protoporfirin IX dan atom besi pusat. Myo-globin merupakan
suatu molekul yang sangat kompak. Besarnya dimensi keseluruhan adalah kurang
dari 45 × 35 × 25 Å, jika urutan sepenuhnya terulur(Gambar. 44). Sekitar 70%
dari rantai utama dilipat menjadi delapan dalam heliks, dan banyak dari sisa
rantai berubah bentuk dan loop antara heliks.


Gambar. 44. Struktur Tiga-Dimensi mioglobin. (A) Model bola-dan-tongkat menunjukkan semua atom nonhidrogen dan
mengungkapkan banyak interaksi antara asam amino. (B) tampilan skematik menunjukkan bahwa protein sebagian besar terdiri dari a-heliks. Gugus heme ditampilkan dalam warna
hitam dan atom besi ditampilkan sebagai bola ungu.
Lipatan dari rantai utama di mioglobin, seperti protein
lainnya, adalah merupakan hal yang kompleks dan tidak memiliki simetri. Untuk keseluruhan
dari rantai polipeptida dari protein ini disebut sebagai struktur tersier.
Sebuah prinsip pemersatu muncul dari distribusi rantai samping. Fakta mencolok bahwa
interior hampir seluruhnya terdiri dari residu nonpolar seperti leusin, valin,
metionin, dan fenilalanin (Gambar, 45). Muatan residu seperti aspartat,
glutamat, lisin, dan arginin yang ada dari bagian dalam mioglobin. Hanya dua
residu histidin yang merupakan residu polar, yang memainkan peran penting dalam
mengikat zat besi dan oksigen. Bagian luar dari mioglobin, pada sisi lain,
terdiri dari residu polar dan nonpolar. Model spacefilling menunjukkan bahwa
ada residu polar dan non polar yang sangat sedikit di dalam ruang kosong. Distribusi
residu polar dan nonpolar mengungkapkan aspek kunci dari arsitektur protein.
Dalam lingkungan berair, lipatan protein didorong oleh kecenderungan kuat dari
residu hidrofobik untuk jauhkan dari air. Ingat bahwa sistem lebih stabil
secara termodinamika ketika gugus-gugus hidrofobik yang terkumpul daripada yang
terurai ke dalam lingkungan berair.

Gambar. 45. Distribusi Asam Amino dalam mioglobin. (A) model ruang mioglobin dengan asam amino hidrofobik yang ditunjukkan dengan warna
kuning, asam amino bermuatan ditampilkan dalam warna biru, dan lain-lain ditampilkan
dalam putih. Permukaan molekul memiliki banyak asam
amino bermuatan, serta
beberapa asam amino hidrofobik. (B) Sebuah
pandangan cross-sectional menunjukkan
bahwa sebagian besar asam amino hidrofobik yang
ditemukan di bagian dalam struktur, sedangkan asam amino bermuatan ditemukan pada permukaan protein.
Rantai polipeptida karena adanya lipatan sehingga rantai
hidrofobik samping yang tersembunyi dan polar, rantai mengarah di permukaan.
Banyak heliks dan untaian-β adalah amphipathic, yaitu α-heliks atau β-untaian memiliki
permukaan hidrofobik, yang menunjuk ke bagian protein, dan permukaan lebih
polar, yang menunjuk ke dalam larutan. keberadaan rantai utama yang menyertai
rantai samping hidrofobik adalah penting juga. Sebuah peptida berpasangan gugus
NH atau CO secara nyata lebih suka air untuk lingkungan yang nonpolar. Rahasia
menyembunyikan segmen rantai utama dalam lingkungan hidrofobik adalah pasangan
semua gugus NH dan CO oleh ikatan hidrogen. Pasangan ini dicapai dalam α-heliks
atau β-sheets. Interaksi van der Waals antara rantai hidrokarbon samping yang padat
juga berkontribusi pada stabilitas protein. Kita sekarang dapat memahami
mengapa set 20 asam amino mengandung beberapa perbedaan dalam ukuran dan
bentuk. Mereka menyediakan palet yang untuk memilih untuk mengisi interior
protein dan dengan demikian memaksimalkan interaksi van der Waals, diaman interaksi
van der Waals membutuhkan kontak intim. Beberapa protein yang span membran
biologis adalah " yang membuktikan pengecualian aturan" mengenai
distribusi asam amino hidrofobik dan hidrofilik seluruh struktur tiga dimensi.
Sebagai contoh, perhatikan porins, protein yang banyak ditemukan dalam membran
luar bakteri (Gambar.46). Hambatan permeabilitas membran yang dibangun sebagian
besar dari rantai alkana yang cukup hidrofobik. Jadi, porins tercakup di luar
sebagian besar dengan residu hidrofobik yang berinteraksi dengan rantai alkana
tetangganya. Sebaliknya, pusat protein mengandung banyak asam amino bermuatan
dan dipol yang mengelilingi saluran berisi air berjalan melalui tengah protein.
Gambar. 46. "Inside Out"
Distribusi Asam amino pada Porin. Bagian luar Porin (yang kontak kelompok hidrofobik dalam membran) sebagian besar ditutupi dengan residu hidrofobik, sedangkan pusat termasuk saluran air yang penuh dilapisi dengan muatan dan polar asam amino.
Jadi, karena fungsi dalam lingkungan hidrofobik porins,
mereka adalah "Inside Out
" relatif terhadap protein yang
berfungsi dalam larutan berair. Beberapa rantai polipeptida lipat menjadi dua
atau lebih daerah yang dapat dihubungkan dengan segmen fleksibel rantai
polipeptida, bukan seperti mutiara pada string. Ini berbentuk daerah bulat unit
kompak, disebut domain, berbagai ukuran dari sekitar 30-400 residu asam amino.
Sebagai contoh, bagian ekstraseluler CD4, protein permukaan sel pada sel-sel
tertentu dari sistem kekebalan tubuh yang human immunodeficiency virus (HIV)
menempel sendiri, terdiri dari empat domain yang sama masing-masing sekitar 100
asam amino (Gambar. 47). Seringkali, protein ditemukan memiliki domain yang
sama bahkan jika mereka secara keseluruhan struktur tersier yang berbeda.
Fungsi
Fungsi

Gambar.
47. Protein Domain. Sel-permukaan protein CD4
terdiri dari empat domain yang sama.
3,5. Struktur Kuarter: Struktur
multi sub-unit dapat dibentuk oleh Rantai Polipeptida.
Pada protein yang mengandung lebih dari satu rantai
polipeptida, protein menunjukkan organisasi struktural tingkat keempat. Setiap
rantai polipeptida protein disebut subunit. Struktur Kuarter mengacu pada tata
ruang dari subunit dan sifat interaksi mereka. Jenis paling sederhana struktur
kuartener adalah dimer, terdiri dari dua subunit identik. Organisasi ini ada
dalam Cro- protein pengikat DNA ditemukan dalam virus bakteri yang disebut l
(Gambar. 48). Struktur kuartener lebih rumit. Lebih dari satu jenis subunit,
sering kali dalam jumlah variabel. Sebagai contoh, hemoglobin manusia, pembawa
oksigen protein dalam darah, terdiri dari dua subunit dari satu jenis (disebut
α) dan dua subunit dari jenis lain (disebut β), seperti yang diilustrasikan
pada Gambar. 49. Dengan demikian, molekul hemoglobin ada sebagai tetramer α-2 β-2.
Perubahan kecil dalam susunan subunit dalam molekul hemoglobin memungkinkan
untuk membawa oksigen dari paru ke jaringan-jaringan dengan efisiensi yang
besar.
Virus terbuat sebagian besar dengan jumlah terbatas
informasi genetik dengan membentuk mantel yang menggunakan jenis yang sama secara
berulang-ulang subunit dalam rangkaian simetris. Mantel rhinovirus, virus yang
menyebabkan flu biasa, termasuk 60 salinan masing-masing dari empat subunit
(Gambar. 50). Subunit datang bersama untuk membentuk shell dengan bentuk hampir
bulat yang membungkus genom virus.
Referensi dan gambar:Jeramy M. Berg, John L. Tymoczko, Lubert Stryer, Biochemistry fifth edition
W.H. FREEMAN and COMPANY
W.H. FREEMAN and COMPANY
No comments:
Post a Comment